研究内容
主に生物の適応進化について理論的に研究しています。
種分化、適応放散、生態系(食物網)進化のシミュレーション
・
ニッチ位置と幅の進化を伴う種分化
・
食物網進化
・
再帰的な適応放散
・
種群全体の適応進化を流体力学的に記述する方程式
Adaptive Dynamics理論(主に進化的分岐条件の多次元拡張)
・
進化的分岐線の条件
・
進化的分岐領域の条件
・
多次元制約下における進化的分岐点の条件
・
潜在分岐条件
・
歪んだ形質空間における進化的分岐条件
・
Adaptive Dynamics理論の適用可能性条件
・
種群全体の適応進化を流体力学的に記述する方程式
統計解析
・
ため池の動物多様性が農薬、コンクリート護岸、外来種、植物減少、水深減少から受ける影響の推定
節足動物の形態形成
・
節足動物の3次元形態形成のための表皮成長制御モデル
動機:
示したこと:
手法:
Adaptive Dynamics理論による解析
無性生殖&有性生殖を仮定した数値計算
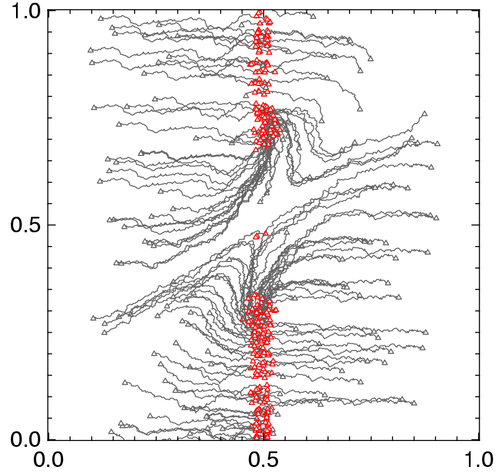
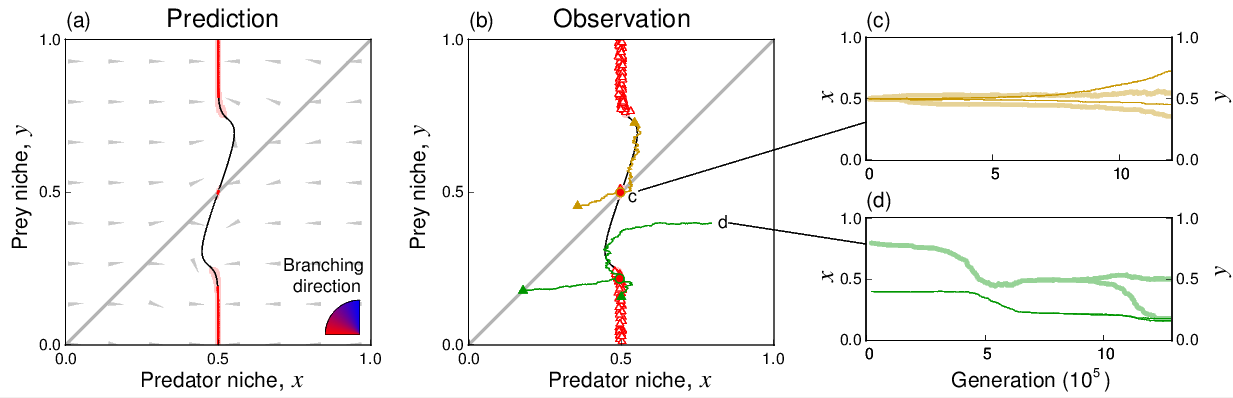
右図の説明:新しい資源(餌)が出現した場合には、元々利用 していた旧資源と新資源の両方を利用するジェネラリストと、旧資源のみを利用するスペシャリストに分化する。(ニッチ位置:主に食べる餌、ニッチ幅:食べる餌の多様性)
Ito H.C., Shimada M. (2007) Niche expansion: Coupled evolutionary branching of niche position and width. Evolutionary Ecology Research vol. 9, 675-695
PDF
動機:
示したこと:
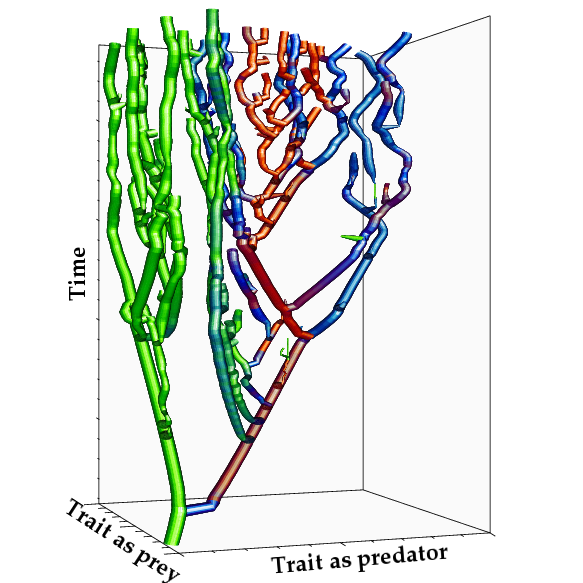
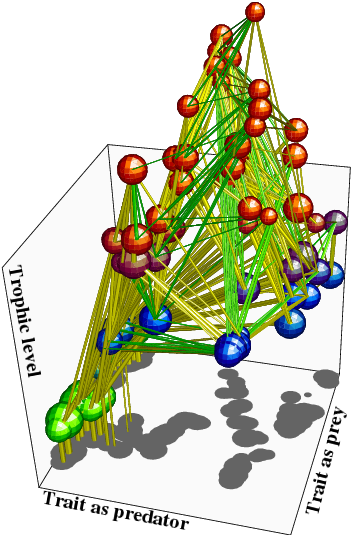
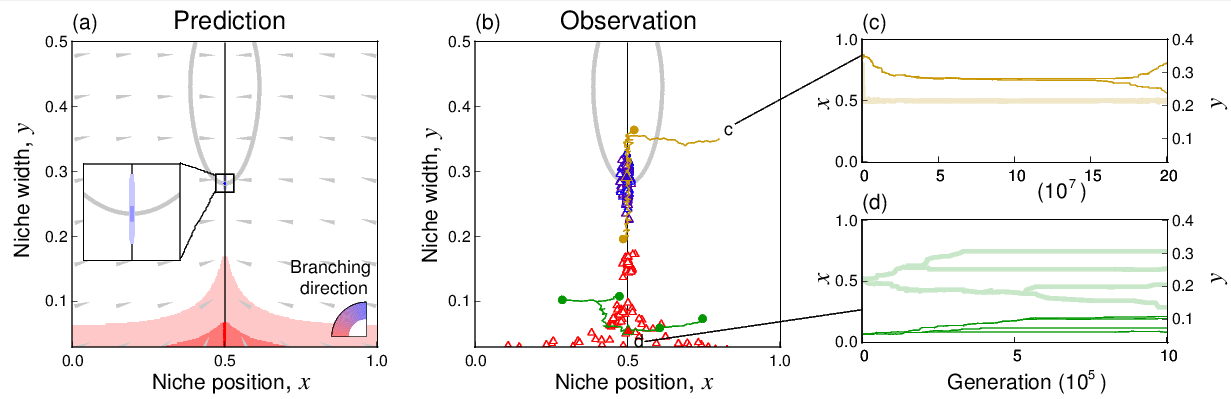
進化的分岐(種分化)を繰り返しにより複雑な食物網 を持つ群集が成長し得る。
群集内の「資源ー資源利用」が理想自由分布に近い状態を維持する場合に、食物網が複雑化する。
手法:
資源競争と捕食被食相互作用を、資源と資源利用パターンの相互作用として統一して記述する数理モデル「拡張資源競争モデル」を考案した(種内レベルから群集レベルまでの個体数動態と進化動態を統一的に解析できる)。
拡張資源競争モデルのAdaptive Dynamics理論による数学的解析。
拡張資源競争モデルにおいて複雑な食物網が成長する進化的経路(中立展開経路)の数学的解析。
拡張資源競争モデルの数値計算(有性生殖を仮定した反応拡散モデル、無性生殖を仮定した確率的モデル)
突然変異を拡散として計算
突然変異を確率論的に計算
Ito H.C., Ikegami T. (2006) Food web formation through recursive evolutionary branching.
Journal of Theoretical Biology vol. 238, pp. 1-10. 2006
Ito H.C., Shimada M.,Ikegami T. (2009) Coevolutionary dynamics of adaptive radiation for food-web
development. Population Ecology vol. 51, 65-81
PDF
動機:
示したこと: 手法:
無性生殖を仮定した数値計算(Oligomorphic stochastic modelという速い計算手法を考案)
有性生殖を仮定した数値計算
無性生殖
無性生殖
有性生殖
Ito H.C., Dieckmann U. (2007) A new mechanism for recurrent adaptive radiations.
American Naturalist vol. 170, pp. E96–E111
PDF
動機:
示したこと:
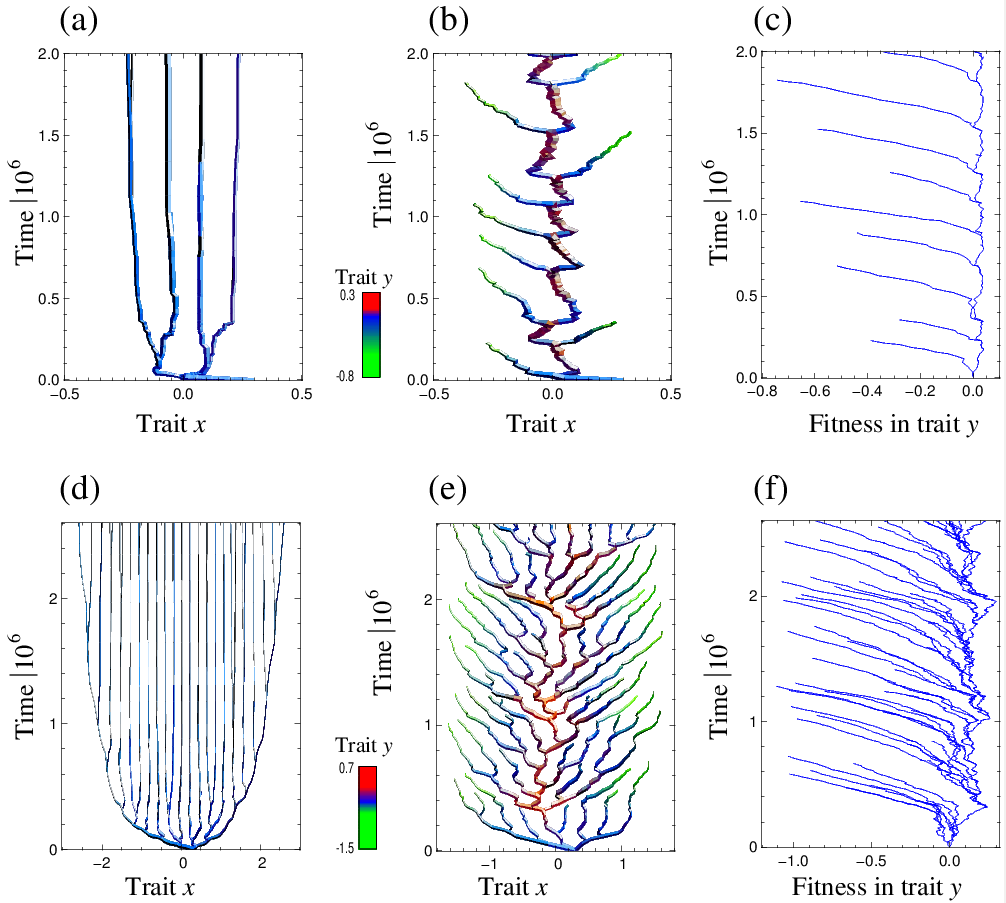
多次元形質空間では、集団がある方向に分断化選択を受けても、別の方向に方向性選択を受けていると、その方向性選択が進化的分岐を抑制し得る。その状況における進化的分岐条件を数学的に導出した。
手法:
各時点での野生型に対して、侵入イベントの確率を最大化する変異型の侵入を想定する「最尤進化経路」を考案。
最尤進化経路において進化的分岐が起きる十分条件(進化的分岐線条件)を導出。
その条件は2次元形質空間では線を成す(進化的分岐線)。
無性生殖集団を仮定した数値計算による進化的分岐線条件の検証。
Ito H.C., Dieckmann U. (2014) Evolutionary branching under slow directional evolution. J. theor. Biol. 360: 290-314
PDF
動機:
示したこと:
進化的分岐線条件を拡張して進化的分岐領域条件を考案した(証明はしていない)。
資源競争モデル(2種類)や拡張資源競争モデルにおいて進化的分岐を数値計算し、進化的分岐線条件と進化的分岐領域条件が有効であることを示した。
手法:
形質空間内の進化的分岐線、進化的分岐領域の数値計算による探索
無性生殖を仮定した数値計算(oligomorphic stochastic model)結果との比較
Ito H.C., Dieckmann U. (2012) Evolutionary branching lines and areas in bivariate trait spaces. Evolutionary Ecology Research vol. 14, pp. 555-582
PDF
動機:
示したこと:
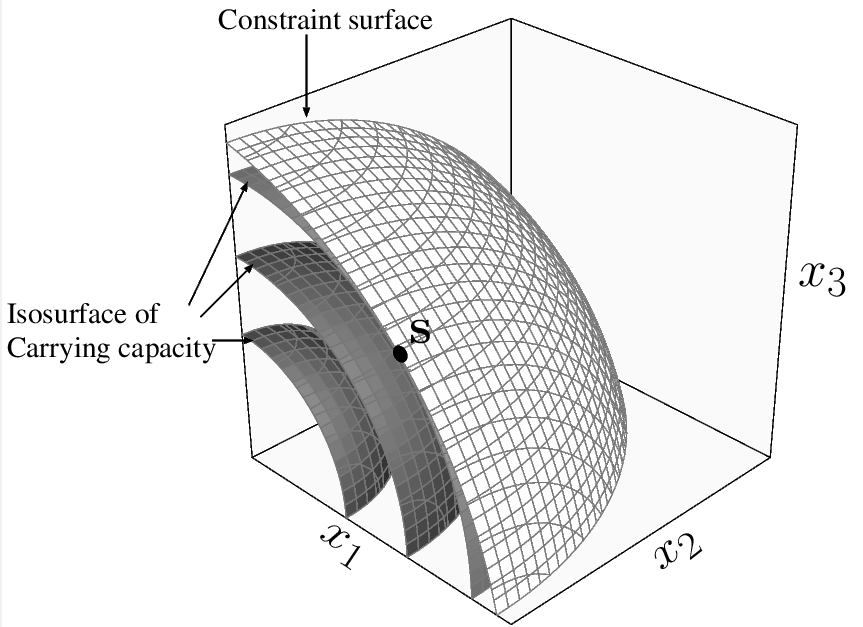
発生学的、遺伝学的、物理的制約などによって、多次元形質空間における進化動態が、より低次元の制約空間に束縛される状況で、任意の次元の制約空間上の進化的分岐点(あるいはその候補)の条件を導出した。
手法:
制約空間上の極値を解析するための強力な手法であるラグランジュ乗数法のラグランジュ関数に相当する「残酷適応度関数」(あるいは「ラグランジュ適応度関数」)を考案。
制約空間上の方向性選択を残酷適応度の1階微分で表現。
制約空間上の進化的分岐点(2次元以上の場合はその候補)の条件を、残酷適応度の1階微分と2階微分で表現。
Ito H., Sasaki A. (2016) Evolutionary branching under multi-dimensional evolutionary constraints. J. theor. Biol. 407: 409-428.
PDF
↑イメージ画像
動機:
示したこと:
個々の数理モデルや相互作用関数に進化的分岐を促す潜在力があるかを吟味できる「潜在分岐条件(branching potential condition)」を導出した。
手法:
制約空間上の進化的分岐条件を抽象化することにより、制約を自由に選べる場合の進化的分岐点である潜在分岐条件を導出。
個々の数理モデルにおいて、固定パラメータや関数を「制約が課せられた形質」として形質空間に加え、潜在分岐条件を適用することにより、そのモデルの核をなす相互作用に進化的分岐を促す潜在力があるかを吟味できる。
Ito H., Sasaki A. (2016) Evolutionary branching under multi-dimensional evolutionary constraints. J. theor. Biol. 407: 409-428.
PDF
動機:
系統的に離れた分類群は同じ選択圧に対して異なる進化的応答を示し得る。
即ち、野生型から生じる突然変異を多次元形質空間における共分散行列(変異行列)で表現すると、その行列は一般に野生型に依存する(形質空間は歪んでいる)。
形質空間の歪みを想定した進化的分岐条件が必要。
示したこと:
任意の次元の歪んだ形質空間における進化的分岐点、線、領域の条件を導出した。
変異の規模が形質(あるいは方向)によって著しく異なる場合に、進化的分岐条件は歪みの影響を受ける。
変異の規模の差異が∞となるとき、進化動態は制約曲面に束縛される。この場合の進化的条件はラグランジュ乗数法による進化的分岐条件(Ito and Sasaki 2016)と一致する。
潜在分岐条件は歪みの影響を受けない。
手法:
形質空間の任意の地点において、その付近を非線形座標変換することで局所的に歪みを消去した後、既存の進化的分岐条件を適用。
Ito H., Sasaki A. (2020) Evolutionary branching in distorted trait spaces. J. theor. Biol. 489: 110152
動機:
Adaptive Dynamics理論の基盤である「アトラクター継承(attractor inheritance)」の証明は限定的なので、拡張する必要がある(アトラクター継承:突然変異の規模が十分に小さければ、変異型の出現が引き起こす適応度地形の変化は僅かであること)。
Adaptive Dynamics理論の適用可能性条件を明確化したい(その条件を満たすあらゆる現象にAD理論を適用できるようになる)。
示したこと:
生態学的に妥当な適応度関数が満たすべき5つの公理を定義。
それらの公理に基づき、AD理論の仮定下で進化的に期待される任意の状況において、アトラクター継承が保証される条件を導出した。
アトラクター継承が保証される任意の進化生態的モデルはロトカ=ヴォルテラ方程式で局所近似できることを示した。
手法:
互いに近縁の表現型をまとめあげておく。
変異体の出現が引き起こす個体数動態の性質を局所リアプノフ関数で解析。
Ito H.C., Dieckmann U., Metz J.A.J. (2020) Lotka-Volterra approximations for evolutionary trait-substitution processes. Journal of Mathematical Biology. DOI
https://doi.org/10.1007/s00285-020-01493-y (オープンアクセス)
示したこと(主に統計解析を担当):
21個のため池において幅広い分類群の動物を調査(爬虫類、魚類、両声類、貝類、甲殻類、昆虫類、線虫類、コケムシ類)。
被説明変数に対して多数の説明変数の候補から有意なものを検出する統計手法を考案。
この手法を適用することにより、BPMC(殺虫剤)の無脊椎動物(特に昆虫)の多様性への有意な負の効果、およびプロベナゾール(殺菌剤)の魚類の多様性への有意な負の効果を検出した。また、コンクリート護岸、外来種(プルーギル)、浮葉植物被度の低さ、水深の浅さの有意な負の効果も検出した。
検出された環境ストレス要因全てによる影響も推定した。
Ito HC, Shiraishi H, Nakagawa M, Takamura N (2020) Combined impact of pesticides and other environmental stressors on animal diversity in irrigation ponds. PLoS ONE 15(7): e0229052.
https://doi.org/10.1371/journal.pone.0229052 (オープンアクセス)
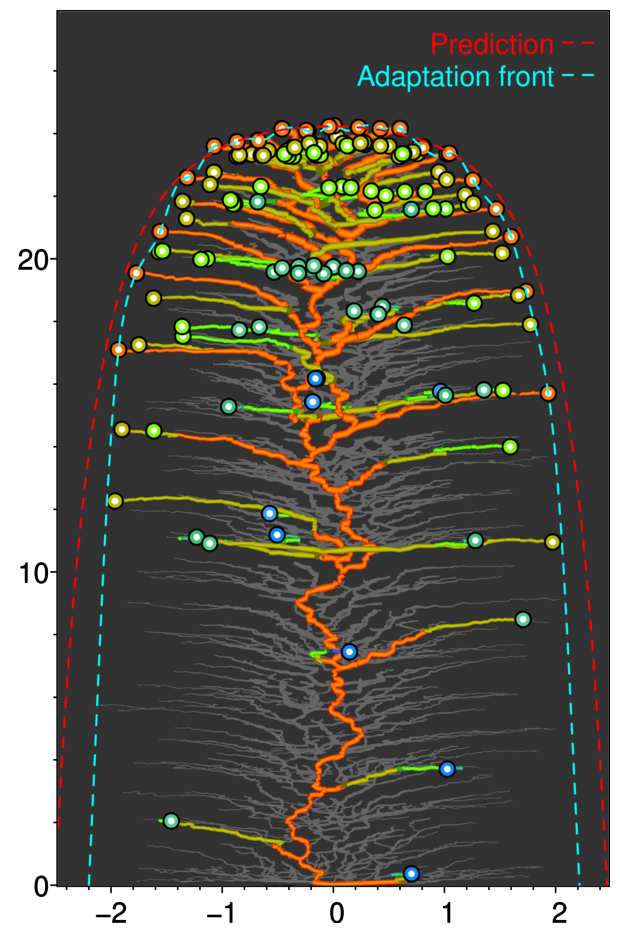
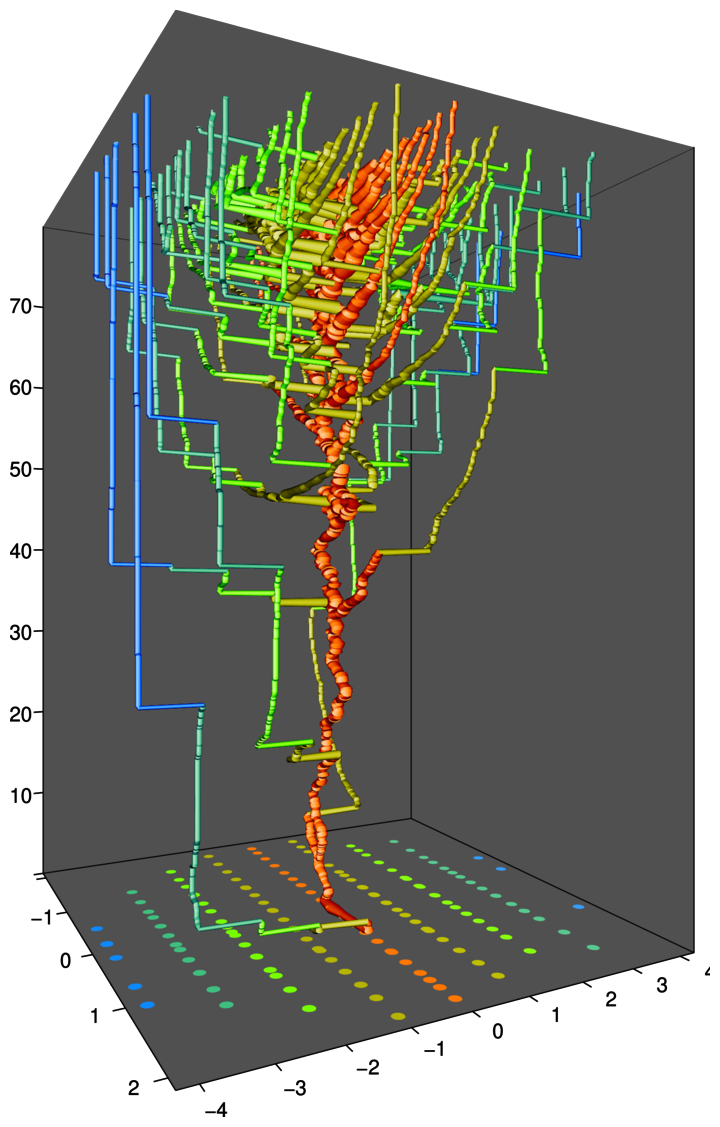
動機:
ニッチの類似限界は分子間力に似てる
進化的革新に起因する放散・絶滅が繰り返されることで、地質学的時間スケールにおける生物の進化的な流れが生じているらしい(チャールズ・ダーウィン、フィリップ・ダーリントン)
示したこと:
各ニッチ位置における平均的な方向進化速度、放散速度、近縁種からの分岐時間、などをニッチ空間における革新速度(主に環境収容力に比例)の分布に基づき予測できる。
辺境のニッチや地理的領域に生息する種の適応進化は著しく遅くなる。その結果、それらの種は同じ時代の最も先進的な種よりも際立って古く、分岐時間も深い「生きた化石」になる。
手法:
進化的革新の繰り返しに起因する再帰的な放散の進化動態において、種群全体の適応進化を流体力学的に記述する方程式を構築した(条件を満たせば任意の形質空間、任意の適応度関数に適用可能)。
数値計算(無性生殖、有性生殖)
[プレスリリース]
適応進化の時計は辺境ほどゆっくり進む: 適応前線方程式が解明する「生物の流れ」と「生きた化石」出現の仕組み
[論文] Ito H. C., Sasaki A. (2023) The adaptation front equation explains innovation-driven taxonomic turnovers and living fossilization. The American Naturalist, 202(6) E163-E180.
https://www.journals.uchicago.edu/doi/10.1086/727046 (オープンアクセス)
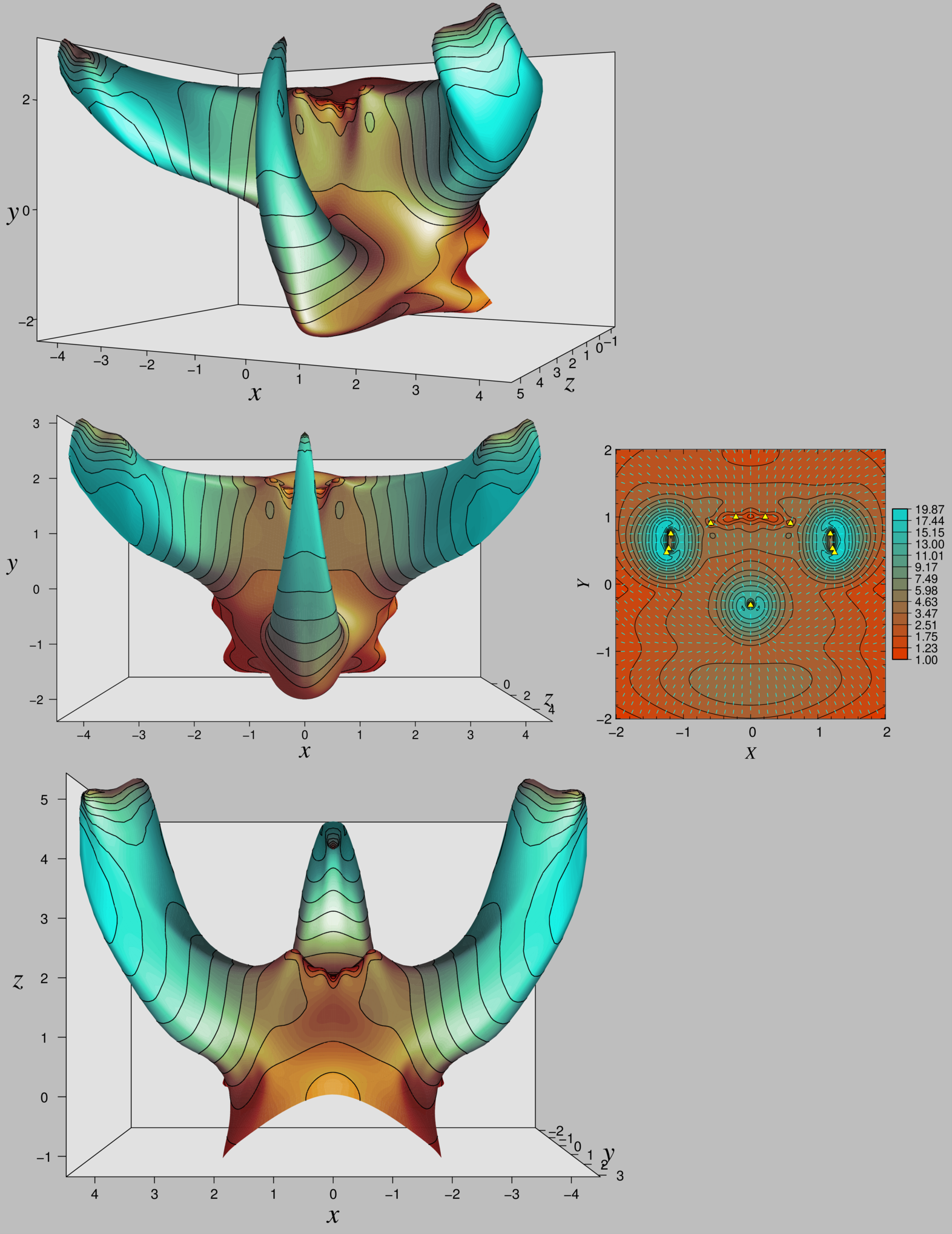
動機:
節足動物は脱皮によって複雑な3次元形態を形成する。
表皮上のモルフォゲン分布が表皮の非一様な成長の制御に関わっている。
示したこと:
表皮上に複数の種類のモルフォゲンを異時的に発現させることで、モジュール的な形態形成(単純な形を組み合わせることによる形態形成)が可能であることを数学的に示した。
節足動物の棘、角、大顎のような形や、カブトムシや糞虫の頭胸部のような形を、平らな膜から作ることができることを示した。
手法:
モルフォゲン分布に基づく表皮の非一様な成長を、表皮の各位置における成長率(成長メトリック)の時間変化として記述する方程式を導出。
成長後の表皮が水圧によって展開される様子を有限要素法やバネ頂点法を計算。
成長した表皮を展開する様子
[プレプリント] Ito H.C., Uchiumi Y. (2024) Growth regulation bringing modularity to morphogenesis of complex three-dimensional exoskeletons. BioRxiv
https://www.biorxiv.org/content/10.1101/2024.05.06.592547v3
[プレプリント] Ito H.C. (2016) Metric dynamics for membrane transformation through regulated cell proliferation. arXiv
https://arxiv.org/abs/1605.01893
 Strange morphs
Strange morphs