1次元のadaptive dynamics理論
前のページ:目次
次のページ:2次元のadaptive dynamics理論
Table of Contents
1 仮定
- 1次元形質空間x
- 突然変異は希で小規模(⇒個体数動態は常にほぼ平衡状態)
2 侵入適応度(invasion fitness)
2.1 定義
侵入適応度は以下のように定義されます(Metz et al., 1996)。
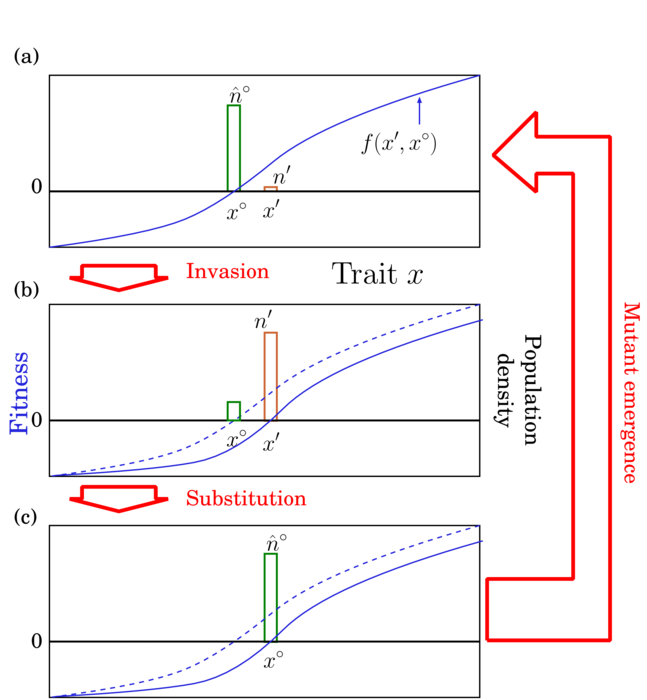
野生型x°に対する変異型野生型x’の侵入適応度f(x’, x°):単型の野生型x°からなる集団の個体数密度n°が平衡個体数密度n̂°にあるとき、出現した変異型x’の初期増加率。
すなわち変異型x’の個体数密度をn’として
すなわち変異型x’の個体数密度をn’として
野生型は平衡密度にあるので常にf(x°, x°) = 0です。
f(x’, x°)は野生型x°の存在下における、形質空間の任意の位置の変異型x’の侵入適応度を与えるという意味で、「野生型x°の存在下における適応度地形」とみなすことができます。この適応度地形はx°に依存して形が変化します。
2.2 形質置換連鎖(trait substitution sequence)
出現した変異型の侵入適応度が正であるとき(f(x’, x°) > 0)、その変異型は野生型に侵入可能であり、侵入した変異型は個体数動態を通じて野生型を駆逐し、変異型が次の平衡状態の野生型となることが多いです(Fig.1)。特別な場合には、変異型と野生型が共存したり、共に絶滅したり、野生型だけが残ることもあります。変異型の侵入が繰り返されることによる野生型の遷移を trait substitution sequence と呼びます(Metz et al. 1996)。ここでは「形質置換連鎖」と呼ぶことにします。
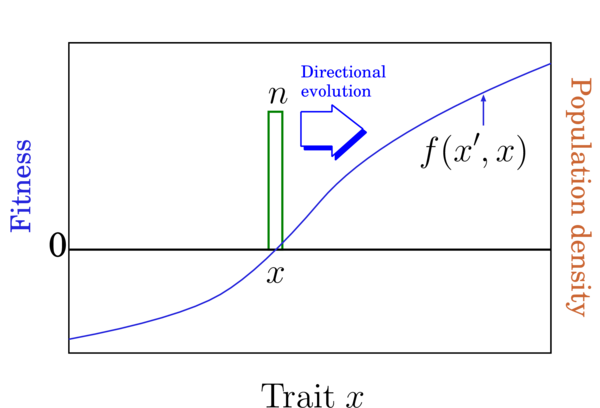
3 方向進化(directional evolution)
形質置換連鎖による集団の方向進化は、野生型の位置の適応度勾配(fitness gradient)
に比例します(Fig.2)。方向進化の期待速度は以下の式により近似的に与えられます(Dieckmann and Law, 1996):
ここでμは突然変異率(1出生あたり)、σμは突然変異の規模(標準偏差)。進化の方向は量的遺伝学におけるLandeの式やPrice方程式と同じですが、速さは異なります。
4 進化的特異点
4.1 進化的特異性(evolutionary singularity)
方向性選択が0になる点を進化的特異点(evolutionarily singular point)と呼びます(Metz et al., 1996)。すなわち、点xが進化的特異点であるとき、
(4) g = g(x) = 0.
4.2 進化的特異点の分類
4.2.1 収束安定性(convergence stability)
進化的特異点xが以下の条件を満たす時、xを収束安定点(Eshel, 1983)と呼びます。
すなわち、x° < xのときg(x°) > 0、x° > xのときg(x°) < 0となるので、集団が単型である限り点xに向かって方向進化し続けます。また、このCは以下のように変形できます。
4.2.2 進化的安定性(evolutionarily stability)
進化的特異点xが以下の条件を満たす時、xを進化的安定点(Maynard Smith and Price, 1973)と呼びます。
この条件は、集団がxに居るときに、周囲の適応度地形が山形になっていることに対応します。
このDが正の場合は谷型の地形に対応します。
4.2.3 相互侵入可能性(mutual invasivility)
進化的特異点xが以下の条件を満たす時、xを相互侵入可能(Prout, 1968)であると表現します。
M, D, Cは − M = D − Cの関係にあります。
5 進化的分岐点(evolutionary branching point)
点xが進化的に不安定な収束安定点であるとき(すなわち以下の3条件を全て満たすとき)、xを進化的分岐点と呼びます(Metz et al., 1996; Geritz et al., 1997)。
(10) C = ⎡⎣(∂g(x°))/(∂x°)⎤⎦x° = x = ⎡⎣(∂2f(x’, x°))/(∂x’2)⎤⎦x’ = x° = x + ⎡⎣(∂2f(x’, x))/(∂x’∂x°)⎤⎦x’ = x° = x < 0. (収束安定性)
- 進化的分岐点の付近に単型(あるいは1山形の表現型分布)の集団があると、その集団は方向進化によって進化的分岐点に到達し、分裂しない限りはそこで分断化選択を受け続けます。
- これらの条件の導出には希な突然変異を仮定しますが、突然変異が稀でないために表現型分布が幅を持つ場合にも有効であることが様々な数値計算によって確かめられています。
6 適用例:資源競争モデル
6.1 個体数動態モデル
ここではロトカ・ヴォルテラの資源競争モデルを考えます。
このモデルはMacArthur-Levins資源競争モデルと呼ばれています。
6.2 侵入適応度
単型x°の集団を考えると、その密度x°の動態は式(12)を用いて以下のようになります。
従ってその平衡密度はn̂° = K(x°)です。変異型x’が出現したとき、その密度の動態は式(12)を用いて以下のようになります。
従って侵入適応度は以下のようになります。
(17) f(x’, x°) = limn’ → 0⎡⎣(1)/(n’)(dn’)/(dt)⎤⎦n° = n̂° = limn’ → 0⎡⎣1 − (n’ + α(x’ − x°)K(x°))/(K(x’))⎤⎦ = 1 − (α(x’ − x°)K(x°))/(K(x’)).
6.3 方向進化
g(x°)
=
⎡⎣(∂f(x’, x°))/(∂x’)⎤⎦x’ = x°
=
− K(x°)⎡⎣(∂α(x’ − x°))/(∂x’) (1)/(K(x’)) − α(x’ − x°)(1)/(K(x’))(∂lnK(x’))/(∂x’)⎤⎦x’ = x°
=
(∂lnK(x°))/(∂x°)
= (∂)/(∂x°)
⎡⎣ − (1)/(2)σ − 2Kx°2⎤⎦ = − σ − 2Kx°
従って、進化的特異点x = 0が存在する。xにおいて、
であるからこの点は収束安定点。さらに、
D
=
⎡⎣(∂2f(x’, x°))/(∂x’2)⎤⎦x’ = x° = 0
=
− K(x°)⎡⎣(∂)/(∂x’)⎡⎣(∂α(x’ − x°))/(∂x’) (1)/(K(x’)) − α(x’ − x°)(1)/(K(x’))(∂lnK(x’))/(∂x’)⎤⎦⎤⎦x’ = x° = 0
=
− K(x°)⎡⎣(∂2α(x’ − x°))/(∂x’2) (1)/(K(x’)) − α(x’ − x°)(1)/(K(x’))(∂2lnK(x’))/(∂x’2)⎤⎦x’ = x° = 0
= −
⎡⎣(∂2α(x’))/(∂x’2)⎤⎦x’ = 0 + ⎡⎣(∂2lnK(x’))/(∂x’2)⎤⎦x’ = 0
=
σ − 2α − σ − 2K = ((σK + σα)(σK − σα))/(σ2ασ2K).
従って、σK > σαのとき、x = 0は進化的分岐点である。(σK < σαのときは収束安定な進化的安定点。)